新城疫
新城疫是由新城疫病毒(Newcastle disease virus,NDV),又称禽副黏病毒-1(Avain paramyxo-virus,PMV-1)引起一种禽的急性、高度传染性疾病,又称亚洲鸡瘟、伪鸡瘟,主要侵害鸡和火鸡,亦可感染其他禽类、鸟类及人类,是世界公认的两大最重要的禽类传染病之一(另一个是高致病性禽流感)。典型新城疫症状为发病急、呼吸困难,头冠紫黑,下痢,泄殖腔出血、坏死,腺胃乳头、腺胃肌胃交界处及十二指肠出血。慢性病例症状比较复杂,常有呼吸道症状或神经等多种症状。由于NDV对感染禽类的高毁灭性,严重影响生产性能,加上世界兽医卫生组织OIE对疫情发生国家的贸易禁运,一旦发生,经济损失巨大,是目前严重危害中国养禽业最主要和最危险的禽病之一。
历史与分布
新城疫1926年首次暴发于印度尼西亚的Java(Kraneveld,1926)和英国的Newcasttie-om-Tyme(Doyle,1927)。尽管此前在中欧已有类似的报告(Halasz,1912)。但真正确定病原的时间是:1927年,经Doyle证明,新城疫与欧洲鸡瘟(禽流感病毒)病原不同,为与鸡瘟区别,根据地名命名为“新城疫”。
在新城疫首次确诊以来的八十年里,世界范围内曾发生过4次NDV大规模的流行。首次大流行源于东南亚和英国,从1926直到六十年代初期,经30多年的时间从印度尼西亚和英国传遍了全世界;第二次大流行始于20世纪60年代后期,从中东迅速传播到大部分国家,此次大流行传播较快,原因是全球养禽业已拓展为国际化和商业化。空中运输禽类及鸟类(笼养鹦鹉)是此次大流行的重要原因。此次大流行造成了严重的经济损失,但却促进了NDV疫苗的研制和免疫预防措施的落实,多数国家加强了对进口笼养鸟的控制;第三次大流行始于70年代中期,发源地和传播尚无定论,由于疫苗的广泛使用,使得引起疫病大流行的病毒更加复杂(Alexander,1997)。第四次大流行始于70年代晚期,起源于中东,传染源是鸽的副黏病毒Ⅰ型(Alexander,1985,1997)。1981年传至欧洲,然后迅速传遍世界各地,一直流传至今。特别是90年代后,该病在欧洲、亚洲许多国家发生,对养禽业造成了巨大的损失。
由于大多数国家对家禽普遍进行强制免疫接种,故对新城疫的地理分布很难评定。目前,只有冰岛、大洋州的少数国家没有新城疫感染,但在1998-2000年,澳大利亚曾爆发过几起新城疫(Kirkland,2000;Westbury,2001)。部分地区如西欧、以色列(A.Panshin,2002)等国也宣布已经消灭了新城疫,但仍有零星新城疫的发生。根据OIE每年发布的疫情公报,2002年发生新城疫的国家和地区为74个,占38.9%;2003年疫区为69个,占37.7%;2004年疫区为68个,占37.9%。2005年和2006年确定有ND临床传染的国家有64个。
我国最早记录该病的发生可追述到1928年的《浙江农业》,1935年,河南有大规模发生,当时认为是“鸡瘟”流行。直到1946年,梁英、马闻天等通过病原分离,证实是NDV,以后在四川、上海、广西等发生。50年代本病在我国各地广泛流行。由于国家及时对该病实行了强制免疫措施,NDV一度得到了较好控制。80年代后,养鸡业在我国蓬勃兴起,饲养规模和数量不断提高,免疫的压力逐渐加大,再加上我国养鸡的主体在农村,设备差,管理水平低,NDV的危害日趋严重,且出现了许多新情况和新问题:80年代发生鸽副黏病毒和鸵鸟新城疫,1997年鹅发生副黏病毒(王永坤,2000)。而传统认为:水禽和其他禽类对NDV抵抗力较强,即使强毒感染,也不表现症状。但是,自97年以后,鹅的发病率逐年升高。此外,自2000年以来,我国山东、江苏、天津、河北等部分养禽发达地区的产蛋鸡群普遍流行一种以临床轻微呼吸道症状、产蛋严重下降(产蛋率通常从90%下降至40%)为主的新城疫,成鸡死亡率较低,但对雏鸡致死率较高。与经典的新城疫(Newcastle disease,ND)不同,发病鸡群通常有较好的HI抗体滴度,一般在28~11(log2),这无疑对于传统的新城疫免疫预防提出了挑战。
病原学
1 分类
ND病毒(NDV)属RNA病毒中的单股负链病毒目、副黏病毒科、副黏病毒亚科、新城疫样病毒(Avulavirus)属,又名禽副粘病毒1型(Avian paramyxovirus 1, APMV-1)。禽副黏病毒目前已鉴定了9个血清型,即APMV-1到APMV-9,NDV属于APMV-1,而其他APMV-2~9血清型的禽副黏病毒中,2、3、6和7型APMV也可侵害家禽并造成经济损失。
围绕新城疫病毒的分类地位,目前尚有争论。De Leeuw (1999)完成NDV
2 形态
新城疫病毒含有单链核糖核酸(RNA),在蛋白质内呈螺旋型对称排列,外部含有类脂性蛋白囊膜。在电子显微镜下,NDV颗粒呈多边性,直径约为100~250nm,有囊膜的病毒粒子一般呈圆形,但因囊膜破损而形态不规则。核衣壳也称基因抗原(G抗原)或核蛋白可溶性抗原(NP抗原),由螺旋状对称直径为17nm的卷曲的核衣壳组成,外面有双层脂质囊膜,病毒囊膜表面覆盖8nm长的纤突(殷震等,1997),分别为HN和F两种糖蛋白,具有能刺激宿主产生抑制红细胞凝集素和病毒中和抗体的抗原成分。NDV的核酸由单股负链RNA组成,全长15186bp,分子量约为5×106道耳顿,占病毒颗粒总质量的0.5%左右。此外,病毒粒子含有约20%~25%(w/w)的来自宿主细胞的脂质和约6%(w/w)的碳水化合物。病毒颗粒在蔗糖中的浮密度为1.18~
3 生物学特性
NDV有许多生物学特性,根据这些特性可与其它黏液病毒相区别。
⑴血凝性(HA):NDV具有凝集红细胞(RBC)的特性,原因是病毒表面血凝素-神经氨酸酶(HN)蛋白具有与红细胞表面相结合的受体。血凝特性及抗血清的特异性在临床上具有诊断意义。NDV可凝集所有两栖动物、爬行动物和禽的红细胞。Winslow(1950)等证明,所有NDV毒株可凝集人和小鼠、豚鼠的红细胞,但凝集牛、山羊、绵羊、猪和马红细胞的能力随毒株而异。一个红细胞凝集单位大约含100000个病毒感染单位。
⑵ 神经氨酸酶活性(NA):神经氨酸酶(NA)是HN蛋白分子的一部分,其功能是可以将病毒从红细胞上洗脱下来,从而使凝集的红细胞缓慢释放。NA还作用于受体位点,使F蛋白能充分接近而发生病毒与细胞膜的融合。
⑶ 细胞融合与溶血活性:NDV和其他副黏病毒引起红细胞溶血或其他细胞融合的机制是相同的。在复制过程中,病毒吸附到受体位点上,病毒囊膜与宿主细胞膜融合(与病毒从细胞中出芽时合胞体的形成相似)。与病毒囊膜融合时,坚硬的红细胞膜常常因破裂而导致溶血。同血凝活性一样,细胞融合与溶血活性可被特异性抗血清所抑制。
⑷其他活性:NDV具有诱导干扰素生成和抗肿瘤等功效。Anan Phuangsab et al(2001)的研究发现,NDV在抗肿瘤免疫,引起细胞凋亡方面有重要作用,使得NDV在抗肿瘤发生、衰老机理等的研究方面备受重视(Karin et al 1994;Jinyang Zeng et al,2002;Sirri et al 2004)。
4 对环境的抵抗力
物理和化学处理均可以破坏NDV的感染性。NDV对热、光等物理因素的抵抗力较强,
NDV对化学消毒药物抵抗力不强,常用的消毒药物,如氢氧化钠、苯酚、福尔马林、二氯异氰尿酸钠、漂白粉等在推荐的使用浓度下5~15min可将病毒灭活;福尔马林、β-丙内酯和酚可破坏病毒的感染性而不能严重损害其免疫原性。
NDV在自然界中的稳定性取决于病毒所处的介质,低温下可长时间,且能使消毒药品无效,故消毒前应先行物理性清洁,而冬季消毒则需在加温条件下进行。
5 毒株分类和致病力
根据血清学中和试验、致病性试验、单克隆抗体、鸡红细胞洗脱率、蚀斑形成的能力和大小、结构多肽、RNA指纹图谱和分子遗传学特性等可将NDV划成不同的种类,但经典的分类方法是:采用多克隆血清进行抗原分型。研究表明:所有NDV分离株均表现为相同的抗原性,因此认为NDV只有一种血清型。但病毒中和试验、琼脂扩散试验和Mcab检测证明不同NDV毒株间存在一定的抗原性差异(Alexander et al,1992)。
根据NDV对鸡和鸡胚毒力的强弱通常将NDV分为三种类型(D.J. Alexander, 1989),即速发型(强毒株,Velogenic)、中发型(中毒力株,Mesogenic)和缓发型(弱毒株,Lentongenic)。不同毒株之间在毒力方面差异较大。
评价NDV毒力致病性的指数主要有3个(见表1),分别是:
MDT(鸡胚平均死亡时间),一般小于60h的为速发型,在60~90h之间的为中发型,大于90h的为缓发型;ICPI(1日龄脑内接种致病指数):一般大于1.60为速发型,1.20~1.60为中发型,小于1.20为缓发型;IVPI(42日龄雏鸡静脉致病指数),一般2.0以上为速发型,2.0以下为中发型或缓发型。需要指出的是:三个指标有时不一定一致,一种指数往往难以确定毒力的强弱,此时应结
表1.新城疫病毒毒株的致病力指数
Table 1 Pathogenicity of NDV isolates
|
毒株 |
致病性 |
ICPIa |
IVPIb |
MDTc |
|
|
无症状肠型 |
0.0 |
0.0 |
>150 |
|
|
无症状肠型 |
0.0 |
0.0 |
>150 |
|
HitchnerB1 |
缓发型 |
0.2 |
0.0 |
120 |
|
F |
缓发型 |
0.25 |
0.0 |
119 |
|
La Sota |
缓发型 |
0.4 |
0.0 |
103 |
|
H |
中发型 |
1.2 |
0.0 |
48 |
|
Mukteswar |
中发型 |
1.4 |
0.0 |
46 |
|
Roakin |
中发型 |
1.45 |
0.0 |
68 |
|
Beaudette C |
中发型 |
1.6 |
1.45 |
62 |
|
|
速发型 |
1.75 |
2.7 |
55 |
|
NY Parrot 70181 |
速发型 |
1.8 |
2.6 |
51 |
|
Italien |
速发型 |
1.85 |
2.8 |
50 |
|
Milano |
速发型 |
1.9 |
2.8 |
50 |
|
Herts, 33/56 |
速发型 |
2.0 |
2.7 |
48 |
a. ICPI,1日龄鸡脑内致病指数
b. IVPI,6周龄鸡静脉内致病指数
c. MDT,鸡胚感染1个最小致死剂量病毒的平均死亡时间(h)
合3种指数综合判定,特别应注重ICPI值的大小。欧洲药典则明确规定:当ICPI≥0.7时,判定NDV为强毒。
6 实验室宿主系统
⑴ 动物 NDV可以试验感染多种禽类、非禽类宿主,鸡是最常用和最易感的自然宿主。根据不同NDV毒株对易感鸡致病力的差异,可将NDV毒株分成三类:即速发型、中发型和缓发型。速发型毒株感染鸡后可引起严重的疫病,出现一种或多种类型的病灶,通常归为死亡。中发型毒株感染鸡后时常症状较轻,病鸡很少死亡;但如将病毒直接引入鸡的中枢神经系统,则发生重病和死亡。而缓发型毒株只可引起轻的或不明显的症状;当用小颗粒对雏鸡进行气雾时,缓发型毒株可引起严重的鸡呼吸道反应。
⑵ 鸡胚 NDV可在鸡胚中繁殖,在SPF鸡胚中易达到高效价,且保持纯净,故SPF鸡胚常用于病毒的分离和鉴定。NDV易在10~12日龄鸡胚的绒毛尿囊膜、尿囊腔中生长,感染的尿囊液能凝集红细胞。强毒株常在24~60h致死鸡胚,缓发型毒株大于100h,中发型则介于中间。
⑶ 细胞培养 NDV可以在多种传代或继代细胞中增殖,包括兔、猪、牛和猴的肾细胞,鸡组织细胞和Hela细胞。最常用的是鸡胚成纤维细胞(CEF)、鸡胚肾和乳仓鼠肾细胞,细胞培养物的病毒浓度通常比鸡胚低1个HA滴度。NDV在大多数细胞系上生长相对鸡胚较差,一般不用于病毒的增殖。NDV在细胞单层上可形成蚀斑,蚀斑的大小与病毒的毒力有关。蚀斑是NDV在细胞上的典型病变。如大多数或全部细胞受到破坏则为透明型,如仅有部分细胞破坏则为混浊型,如无病变,则仅为细胞对染料的渗透,表现为红色型。蚀斑在接种病毒后2~6天内形成。一般实在接种后96h进行观察。在大多数细胞培养物中,如果覆盖层不加Mg2+和DEAE或胰酶,仅速发型和中发型NDV毒株能在鸡胚细胞上形成蚀斑。慢性感染的细胞培养物常不显现蚀斑。病原分离时,可利用蚀斑技术,连续传三代建立NDV的克隆,进而纯化病毒。
流行病学
NDV传统仅对鸡、火鸡等表现较高的死亡率,其他禽类很少感染,而目前宿主范围明显扩大,已经有鹅,企鹅,鸭,鹦鹉,鸽子,鸬鹚,驼鸟,野鸡,孔雀,秃鹫,鸱鸪,朱鹗等发病的报道,能自然或人工感染的鸟类超过250余种。
最引人关注的是鹅的NDV感染,在1997年以前,水禽尤其是鹅感染NDV发病的报道较少。即使人工攻毒,水禽的发病率仍较低。但自1997年以来,在江苏、山东、浙江、四川等地陆续分离出具有高发病率和死亡率的鹅NDV,造成了严重的经济损失(王永坤,2000)。研究表明,家鹅比家鸭易感,番鸭又比其他品种的鸭易感。人也可感染,引起急性结膜炎。最近有猪感染NDV的报道。
在自然条件下,病毒主要经呼吸道或眼结膜感染,也可经消化道感染。病鸡的分泌物、排泄物和尸体是病毒的主要传染媒介。病鸡在出现症状前24小时,分泌物和粪便中就含有大量病毒。病愈鸡在症状消失后1周即停止排毒,个别鸡的排毒期可达一个月以上。病鸡经呼吸道和消化道可向外排出大量病毒,污染周围环境而成为传染源。病毒通过被病毒污染的垫料、饲料、饮水、水槽和用具传播给健康鸡。带毒鸡和携带病毒的人员流动对传播本病起到重要的作用。
本病无季节性,但以春秋发病最多。鸡舍通风不良,氨味过大,感染禽流感、法氏囊等疫病,各种应激等均是爆发本病的诱因。当一个鸡群发病死亡较高,首先应考虑是否是新城疫。近些年来,非典型新城疫发生较多。其流行特点是:多发生在用疫苗免疫过的鸡群,以二免前后发生较多。雏鸡和成年鸡的发病率和死亡率都不高。非典型新城疫容易成为持久性感染,鸡群中由于陆续出现病鸡,长期不断排出病毒,干扰疫苗免疫效果,造成疫病的连续传播,在鸡群中绵延不断,特别在在大群舍养鸡中更是这样。
传统认为NDV不能经卵发生垂直传播,因为病母鸡所产的蛋,在孵化的头4~5天,胚胎会因感染而死亡。但最近不少报道从种蛋中分离出NDV。
值得一提的是鹅的新城疫感染。不同日龄的鹅具有感染性,发病最小的为3日龄,最大的300日龄,日龄愈小,发病率和死亡率愈高,随着日龄增长,发病和死亡下降,2周龄以内的雏鹅,发病率和死亡率可达100%。根据不同鹅群的调查,发病率40%-100%,平均60%左右,死亡率30%-100%,平均为40%左右。同群饲养的鸡通常在鹅发病后2-3天也感染发病。鸭感染发病的报道虽少,但有增加的趋势。
临床症状
鸡新城疫临床潜伏期长短,与病毒毒力和侵入鸡体内的病毒量以及个体的抵抗力有关,人工感染为2~5天,自然感染时潜伏期一般为3~5天,毒力弱的可延至20天。根据临诊表现和病程长短,病鸡的症状可分为最急性、急性、亚急性或慢性以及亚临床和非典型四种类型。
1.最急性型 此型多见于流行初期,突然发病,常无特征性症状而突然死亡,雏鸡和中雏为多见。
2.急性型 在发病初期体温升高,一般可达43~
3.亚急性或慢性型 初期症状与急性相似,不久后症状逐渐减轻,同时出现神经症状。病鸡翅膀和腿麻痹,表现站立不稳,头颈向后或向一侧扭曲。有的病鸡貌视正常,但受到惊动时,突然伏地旋转,动作失调,反复发作,终于瘫痪或半瘫痪。有的病鸡因吃食受到影响渐渐消瘦,终归死亡。此型多发生于流行后期的成年鸡,病死率较低。
4.免疫鸡群的非典型或亚临床症状 近几年来,在免疫鸡群中发生往往亚临床症状或非典型新城疫,发病率和病死率较低,一般在5%~45%。主要表现为呼吸道和神经症状,当呼吸系统症状减轻趋于康复时,少数病鸡遗留头颈扭曲。产蛋鸡(20周龄以上的)感染新城疫,主要出现产蛋下降,严重者产蛋下降90%左右,常有呼吸道症状,病死率较低。这种类型的新城疫无论从临床症状和病理变化上均不易诊断为新城疫,但当进行病原分离时,却往往能分离到高度致病性的新城疫病毒。这一类型的新城疫在临床上有以下特点。
⑴ 产蛋鸡:发病鸡群通常处于产蛋高峰,临床上突然出现轻微的呼吸道症状,产蛋率一般下降40%~60%,种蛋受精率和孵化率也随之下降,其他无明显异常,也可能有较明显的呼吸道症状和死亡数增加,但很少见有消化道明显出血的病例。抗体跟踪检测发现:鸡群未发病前ND的平均抗体水平为28.5,而发病后ND抗体可飙升至212~15。
⑵ 非产蛋鸡:症状不一,可能出现不同程度的呼吸道症状,有的鸡群表现为少数鸡摇头和咳嗽,有的只能在安静时才能听到轻微的呼吸道啰音,而有的鸡群则可出现明显的呼吸困难,喉头充血、出血,甚至咳血,但死亡率不高,一般不超过30%。
火鸡、鹅、鸽、鸵鸟、珍珠鸡、雉鸡、鹌鹑和鸭等感染新城疫后的临床症状与鸡基本相同,只是程度上有差异而已。
以下以鹅新城疫为例介绍:潜伏期一般为3~5天,病程为2~5天。患病鹅初期拉灰白色细粪,病情加重后粪便呈水样,带暗红色、黄色、绿色或墨绿色。病鹅精神萎顿,眼有分泌物,常蹲地,少食或拒食,饮水增加,体重减轻,行动无力,浮在水面。后期部分鹅出现扭颈、转圈等神经症状。有的发病鹅有呼吸道症状。
病理变化
1.剖检病变
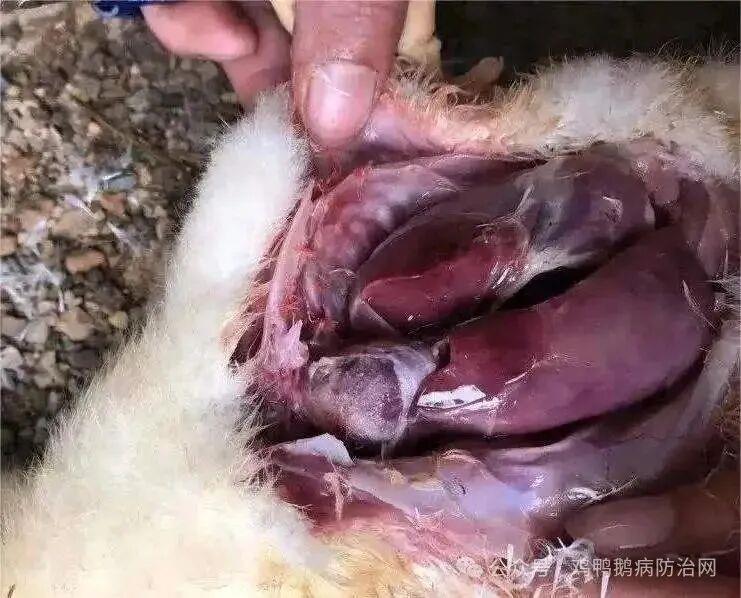
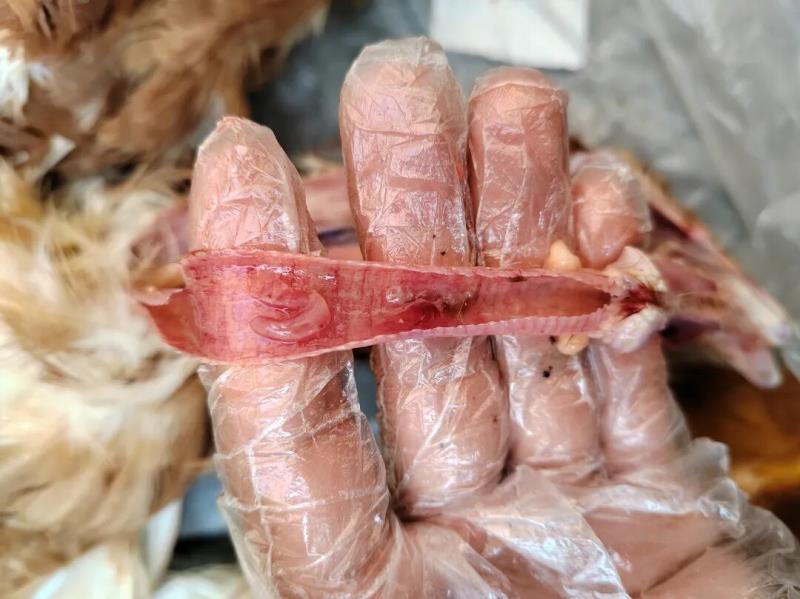
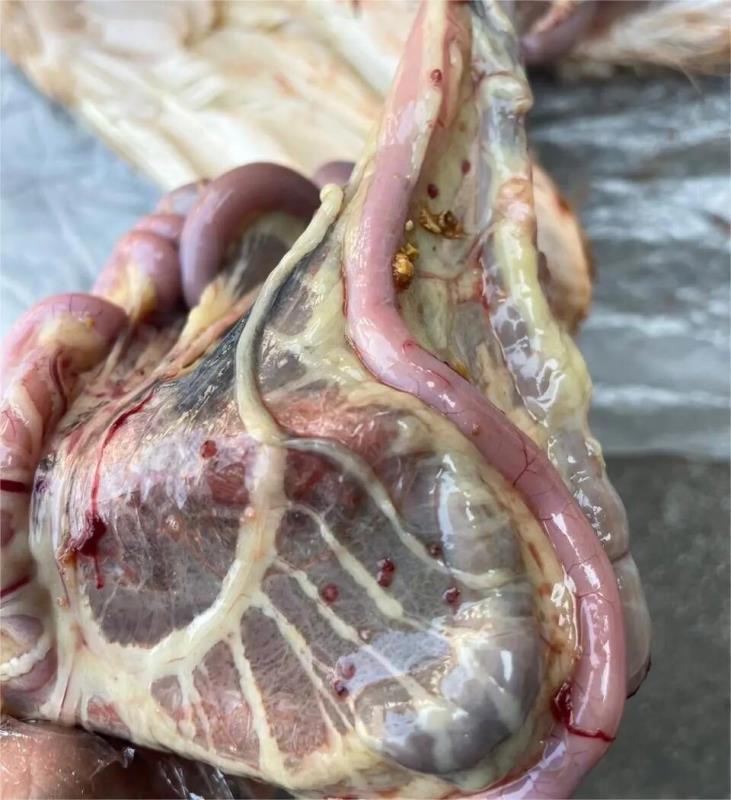
典型的新城疫病变:病死鸡鸡冠和肉髯紫黑色;口腔内充满粘液,嗉囊内充满硬结饲料或气体和液体;泄殖腔充血、出血、坏死、糜烂,带有粪污;腺胃乳头出血,腺胃与肌胃交界处及腺胃与食道交界处呈带状出血,肌胃角质膜下出血,有时还见有溃疡灶;十二指肠以至整个肠道粘膜充血、出血;喉气管黏膜充血、出血;心冠沟脂肪出血;输卵管充血、水肿,其他组织无特征性病变。
病死鹅皮肤淤血,部分病例皮下有胶样浸润。患鹅脾脏肿大、淤血,有大小不等灰白色坏死灶。肠道黏膜出血、坏死、溃疡。部分病例的腺胃及肌胃黏膜充血出血。心肌变性,有的心包积液。出现神经症状的病例。出现脑充血、出血和水肿。
非典型新城疫病变:主要来自有免疫背景的鸡群,其体内含有NDV的体液抗体,这些鸡发病后,往往看不到ND的典型病变。常见的病变是心冠脂肪的针尖大出血点,腺胃肿胀和小肠的卡他性炎症,盲肠扁桃体普遍有出血,泄殖腔也多有出血点。气管内黏液增多,气囊混浊并有干酪物。成鸡免疫后感染NDV时,也往往不出现ND的典型病变,较常见的病变是盲肠扁桃体出血与泄殖腔有出血点。必须多剖检一些病例,才能发现腺胃乳头和肌胃角膜下、十二指肠黏膜轻度出血等ND典型病变。
2.组织学病变
患病鸡大部分脏器均可见充血、出血、水肿、坏死等变化,有大量淋巴细胞浸润。在急性病例中各器官,特别是肝脏有网状组织细胞增生,脾的网状细胞内有包涵体。在肝中有细小的坏死病灶。肠内出血性坏死病变在淋巴集结中形成。神经系统一般不出现脑炎,若出现时则为神经胶质病变,神经原变性和血管周围淋巴细胞侵润以及内皮细胞肥大,病变分布在中枢神经系统—神经索、延髓、中脑和小脑。生殖系统有卵巢炎和输卵管炎。
患病鹅组织病理变化主要在脾脏、胰腺和心肌。脾髓淤血,实质淋巴细胞明显减少,脾小体几乎完全消失,坏死灶大小不一,很多融合一片。胰腺实质腺泡上皮变性,有大小不一的坏死灶。心肌纤维广泛颗粒变性,可见很多心肌纤维发生坏死,断裂或崩解。
诊断
由于典型新城疫的症状和病变与高致病力禽流感十分相似,因此仅凭临床症状与病理变化很难做出准确的诊断。必须进行病毒分离鉴定和其他的实验室诊断才能确诊。
1.病原学检查
标本采取 用感染后3~5天的病禽的组织器官、体液和分泌物内,均易分离获得病毒。其中以肺、脾、脑内的病毒含量最高,骨髓内含毒时间最长。需向实验室寄送标本时,最好割取鸡头,用油纸包扎并于冷藏条件下寄去,这在典型新城疫以及呈现神经症状的病鸡,常易分离到病毒,也可用棉拭子从气管和泄殖腔取样。接种过疫苗的鸡发生感染时,采取标本的时间应推迟至感染后的6~14天。
2.病原分离和鉴定
①鸡胚 最好选用9~11日龄的SPF鸡胚。将病料作尿囊腔接种,置于37~
②动物回归试验 接种易感幼鸡,观察其临床发病和病理剖检情况。
③细胞培养 分离新城疫病毒,可以采用鸡胚成纤维细胞。必须指出,某些新城疫毒株不能在细胞培养物上产生规律性细胞病变,血凝滴度也远比鸡胚液低。分离病毒以后,即可应用血凝抑制试验予以鉴定。
3.特异抗体的检测:主要用于流行病毒分病学调查和疫情回顾性诊断。
应用高滴度的感染鸡胚尿囊液(血凝滴度1:320 以上)作为抗原,稀释成4个血凝单位的稀释液后,加入等量倍比稀释的被检血清,按常量法、微量法、平板法或全血法进行操作。由于鸡群普遍接受疫苗接种,难对单份血清的血凝抑制滴度进行评价,因此,应以双份血清中恢复期血清滴度比急性期血清高4×以上者作为阳性指标。
4.病毒的鉴定
因禽流感病毒、减蛋综合症病毒等其他一些病毒也具有凝集鸡红细胞的活性,所以在HA呈阳性后,还必须用新城疫抗血清对分离的病毒进行红细胞凝集抑制试验(HI试验),最好同时分别用新城疫、禽流感和减蛋综合症的单因子抗血清与分离物作HI实验。如果新城疫抗血清能抑制被分离物的HA活性,而其他血清不能将其抑制,则被分离的病毒为新城疫病毒;如病毒的HA活性分别能被新城疫及禽流感的抗血清所抑制,则提示分离物在感染了新城疫的同时还混合感染了禽流感。
由于养禽业普遍应用弱毒疫苗,且自然界中存在着大量不同毒力的毒株,禽类中又有比较严重的带毒现象,故从动物体内分离获得新城疫病毒,并不必然证明这次疫情就是新城疫。因此,必须测定新分离毒株的毒力,以便建立最后诊断。通常应用最小致死量致死鸡胚时间平均值(MDT)、脑内接种雏鸡致病指数(ICPI)以及静脉接种幼鸡致病指数(IVPI)等三种方法测定病毒毒力。
具体的方法是:
(1) 病毒对1日龄雏鸡脑内接种的致病指数(ICPI)
检测方法为:取血凝实验(HA)滴度24以上的感染了NDV的新鲜尿囊液,用等渗无菌盐水做10倍稀释,不添加抗生素等物质。脑内接种出壳24~40h之间的SPF雏鸡,共接种10只,每只接种0.05ml。每24h观察1次,共观察8d。每天观察应给鸡打分,正常鸡记作0,病鸡记录1,死鸡记作2(每只死鸡在其死后的每日观察中仍记2)。ICPI是每只鸡8d内所有每次观察数值的平均数。
最强毒力的NDV的ICPI接近最大值2.0,而温和型毒株的接近0。各国在确定强毒、中等毒力和弱毒株的ICPI值范围上不尽相同。
(2) 静脉致病指数(IVPI)
检测方法为:取HA滴度高于24的已感染NDV的新鲜尿囊液,用灭菌等渗盐水做1:10稀释。由静脉接种6周龄的SPF小鸡,接种10只,每只鸡接种0.1ml。每天观察1次,连续10d,每次观察并记分,正常记作0,病鸡记作1,瘫痪鸡或出现其他神经症状记作2,死亡鸡记作3(每只死鸡在其死后的每日观察中仍记3)。IVPI是指10d内每次观察每只鸡的记录的平均值。
温和型毒株和一些中等毒力NDV株的IVPI值为0,而强毒力株可达到3.0。各国在确定强毒、中等毒力和弱毒株的IVPI值范围上不尽相同。
(3) 鸡胚平均致死时间(MDT)
检测方法为:将新鲜的感染尿囊液用生理盐水连续10倍递增稀释至10-6~10-9。每个稀释浓度接种5个9~10日龄的SPF鸡胚,每个鸡胚接种0.1ml,随后置于
5.其他诊断方法 包括血清中和试验、免疫荧光抗体技术、ELISA、单克隆抗体技术、核酸探针等分子生物学技术等。由于目前鸡群已普遍接种新城疫的疫苗,血清学方法如未能区分抗体来自疫苗还是野外病毒之前,必须比较感染前后的抗体滴度是否有明显上升,才具有诊断意义。
6.鉴别诊断
①高致病AI 两种病毒对各种品种和年龄家禽均具有高度致病性,死亡率高。但新城疫潜伏期较长,有特殊的呼吸罗音,绿色下痢,食欲减退,后期出现神经症状。在病理变化上,高致病性AI以全身器官出血为特征,典型新城疫则表现上呼吸道和消化道炎症,肠道“钮扣状”溃疡和腺胃乳头出血是特有病变。目前养殖条件下,新城疫疫苗免疫鸡群很少发生典型新城疫。
②禽霍乱 禽霍乱病原为巴氏杆菌,病程急,死亡快。病死禽肝脏有散在性或弥漫性针头大小的坏死灶,广谱抗生素和磺胺药对禽霍乱有紧急预防和治疗作用,而抗生素对新城疫治疗无效。
③低致病性AI 低致病性AI对产蛋鸡影响较大,可使产蛋下降30%-70%,甚至停产。剖检见气管粘膜充血出血,有黄色干酪样物阻塞,特别是与大肠杆菌混合感染时可导致严重的气囊炎,气囊膜增厚,其上附有多量的黄色干酪样物。产蛋鸡输卵管炎,常见蛋清样分泌物或干酪样栓塞。非典型新城疫成鸡主要表现为产蛋量下降,幅度为10%-30%。青年鸡发病神经症状较突出。剖检喉头及气管充血、出血,有少数鸡可见腺胃乳头有少量出血和肠道“钮扣状”溃疡。
④传染性支气管炎 传染性支气管炎仅发生于鸡,幼雏发病严重,6周龄以上和成年鸡发病后症状较轻,病死率一般为0.5-1%。产蛋鸡仅表现产蛋减少,产畸形蛋、小蛋,薄壳蛋,蛋白稀薄如水。剖检死亡鸡在支气管有干酪样栓子。
免疫与防制
新城疫是OIE一类疾病,我国动物一类疫病,危害极大,国际上曾试图通过统一的法规来防止新城疫的发生,但由于不同国家经济水平不同,很难达成共识。为防止病毒的传入,各国均制定了严格的检疫制度。防止带毒的禽及其产品流入。对于新城疫无有效的治疗方法,发达国家多采用扑杀的方法控制ND的发生,我国主要采取免疫预防与扑杀相结合的综合防制措施。
(一)免疫的机理
NDV侵入机体后主要引起败血症,可诱导产生黏膜抗体、中和抗体和血凝抑制(HI)抗体等。病毒中和抗体能有效地阻碍病毒对鸡、鸡胚和细胞培养致死性感染能力。
体液免疫
在ND免疫反应中,体液免疫起着十分重要的作用。鸡在初次感染NDV后,通常于6~10天即可测出HI和中和抗体,二者的高峰反应一般在第3~4周出现,在4个月内HI抗体的滴度能维持一定水平,到8个月时开始下降直至消失。HI抗体同中和抗体一样,可保护鸡、鸡胚和培养细胞免受NDV的感染,二者具有密切相关性。通常用HI抗体作为评价疫苗免疫接种后抗体水平好坏的标志。不同疫苗毒株刺激产生抗体高低的能力不一。接种ND活疫苗和灭活苗均能产生体液抗体。
细胞免疫 细胞免疫在新城疫的免疫和感染中具有重要作用,但细胞免疫不能完全抵抗鸡新城疫病毒入侵,它能与体液免疫协同发挥作用(刘胜旺等,1998)。感染NDV的最初免疫反应是受细胞调理的,并可于免疫活疫苗后2~3天很快检出。这大概可以解释:为什么家禽在免疫后可测出抗体之前就具有一定的早期保护性。Ghumman(1976)等发现,接种ND活疫苗和灭活苗后2~3天,就能检测到淋巴细胞内DNA合成增加。Agrawal(1991)等用白细胞游走抑制试验来评价接种ND疫苗后的细胞免疫反应,发现接种后确实存在细胞免疫反应,但细胞免疫反应的水平和抗体水平之间不存在平行关系,细胞免疫的作用不如体液免疫。
局部免疫 主要是黏膜免疫。黏膜特异性免疫系统是由黏膜相关淋巴组织,包括肠相关淋巴组织和支气管相关淋巴组织组成的免疫系统。包括肠道黏膜集合淋巴结、消化道、呼吸道和泌尿生殖道黏膜下层的淋巴小结和弥散淋巴组织,含有丰富的B细胞,T细胞和巨嗜细胞等。NDV感染的主要门户是消化道和呼吸道。因此,在局部建立起有效的黏膜免疫对于NDV的控制意义重大。黏膜免疫参与的主要成分是分泌型IgA(SIgA)和一类能下调全身性免疫应答的效应性T细胞,以及黏膜定向细胞运输系统。SIgA对机体呼吸道、消化道等局部黏膜免疫具有相当重要的作用,特别是对于一些经黏膜途径感染的病原微生物,动物机体这种黏膜免疫功能就显得十分重要,若动物机体呼吸道、消化道分泌液中存在这些病原微生物的相应的SIgA,则可抵抗其感染,因此,SIgA是机体蛋白膜免疫的一道“屏障”。 SIgA细胞的产生与抗原直接剌激黏膜有关。在传染病的预防接种中,经滴鼻、点眼、饮水和喷雾途径免疫弱毒活疫苗均可产生SIgA而建立相应的黏膜免疫力。活疫苗经过饮水、滴鼻和喷雾等接种途径,能够刺激黏膜免疫,诱导局部合成和分泌IgA抗体。Herbert(1994)认为,灭活病毒接种后不出现IgA和IgM,仅有高水平的循环抗体IgG,故灭活疫苗不能产生黏膜免疫。正因为如此,机体容易缺乏局部的黏膜免疫,不能有效地抵抗外界野毒的侵袭。因此,在进行免疫时,应以弱毒活苗作基础,配合油乳剂灭活苗。
(二)疫苗的免疫
应用疫苗对易感鸡进行免疫接种而获得免疫力,抵抗外界NDV强毒的攻击,这就是通过人工免疫而获得主动免疫。疫苗免疫是我国控制NDV强毒的基本国策。
1.疫苗种类 新城疫的疫苗有两大类,一为活疫苗,二是死疫苗。
活疫苗 弱毒疫苗种类较多,不同的种类其免疫性能不一样,免疫的方法也不完全相同。一般分为两大类:中发型和缓发型疫苗。中发型:代表毒株为Ⅰ系,其致病力较强,对雏鸡可能致死,适合2月龄以上的鸡,但其免疫原性好,产生抗体速度较快,3~4天即可产生免疫力,维持时间较其它活疫苗长,常使用肌肉注射或刺种,适用于有基础免疫的鸡群和成年禽接种,常用于加强免疫。克隆Ⅰ系来源于Ⅰ系疫苗,免疫原性相当,但毒力较弱,是目前Ⅰ系的理想“替代品”。
缓发型:代表毒株为B1、LaSota株等毒株,毒力相对较弱。Ⅱ系疫苗毒力弱、安全性好,主要用于雏鸡免疫,但抗体维持时间较短,另外母源抗体可影响其免疫。Ⅲ系疫苗特性与Ⅱ系非常相似。Ⅳ系疫苗是使用最广泛的新城疫疫苗,毒力比Ⅱ系、Ⅲ系高,免疫原性好,维持时间较长。另外常用的有V4和Clone 30,其中有报导V4的粘膜免疫效果较好,可在局部如气管抵抗新城疫病毒的入侵;Clone 30与Ⅳ系很相似。接种方法常用滴鼻、点眼和饮水等。对于弱毒疫苗在选择要注意其对黏膜的亲嗜性,传统的疫苗如:Lasota 系、C30、Ⅱ系、F系等,属于对呼吸道黏膜亲嗜性较强的疫苗,可以刺激呼吸道黏膜免疫的效应。而有些疫苗如:V4对消化道黏膜有特殊的亲嗜性,也可以产生呼吸道黏膜的免疫效应。
死疫苗 即“灭活疫苗”,来源于感染性NDV尿囊液,用β丙内酯或福尔马林杀灭病毒,与佐剂混合。常用矿物油作为佐剂生产油乳剂灭活苗。灭活苗不像活疫苗容易被母源抗体中和,接种后产生抗体比活疫苗慢,但抗体高峰持续的时间比活疫苗长。Levy等(1975)研究表明,2周龄的试验鸡经肌肉或颈部皮下接种灭活疫苗,免疫后第14天HI抗体平均效价为1:40,免疫后20天抗体达到高峰,效价1:496,抗体高峰维持3个月才开始下降,直至第5~6月仍达1:100左右。
2. 影响疫苗免疫效果的因素
要想产生良好的免疫效果,不仅要有好的疫苗,更要有科学的免疫程序。但免疫程序不是一成不变的,而是要根据各鸡场、地区不同情况而异。影响免疫效果的主要因素有:
①母源抗体:母源抗体直接影响疫苗的免疫效果。若母源抗体低,很容易导致雏鸡的早期感染;若高,则可中和疫苗毒,导致免疫效果差,抗体水平低。雏鸡母源抗体效价的下降有一定规律,每隔4.5天下降一半,如3日龄时HI效价为1:128,则7.5日龄时下降为1:64,12日龄时下降为1:32,据此可计算出雏鸡在12日龄后可进行免疫接种。所以对雏鸡测一次HI效价就可计算出它在几日龄时适于作免疫接种。母源抗体的HI效价越低,其免疫效果越好,一般要求在1:32以下时进行免疫接种。但若处在ND严重污染地区则需在1:64时即进行免疫接种,1:128以上时对强毒攻击有抵抗力,同时对疫苗接种基本上不起反应,所以母源抗体在1:128以上时暂不要用苗。当前ND免疫失败的原因多数是由于母源抗体过高所产生的干扰所致,应结合母源抗体的抗体消长规律进行合理免疫。
②疫苗接种途径的影响:疫苗的接种途径与免疫效果的高低有直接的关系。当鸡作个体接种时,以皮下或肌肉注射的效果为最好;滴眼次之,滴鼻又次之。当鸡作群体接种时,气雾的免疫效果最好,饮水次之,喷粉和饲喂(V4弱毒株可混于饲料中)应用较少,效果也较差。
③ND疫苗种类和次数:目前应用较广泛的是油佐剂灭活苗。油佐剂灭活苗对鸡安全、免疫效果好,雏鸡免疫一次能较活苗的免疫效力维持更长一些时间,灭活苗的特点是免疫持续时间长、抗体水平高,至少有3个月的免疫期,但缺点是成本较高、操作麻烦、产生抗体速度较慢,灭活苗的佐剂有氢氧化铝、植物油、矿物油及蜂胶等,使用佐剂可延长免疫期、增强免疫效果。实际生产中常同时使用活苗与灭活苗,既可较快产生抗体,又可维持较长时间的高水平抗体。无论是弱毒苗和灭活苗,二次免疫均能出现强烈的免疫应答反应,抗体水平有较显著提高。对于已接种过活疫苗的鸡,再接种灭活苗,可显著提高抗体水平。杨增岐等(1997)发现,在较高母源抗体的情况下,单用活疫苗给雏鸡免疫,效果不理想,而用活疫苗和灭活苗同时接种,可产生较高水平的抗体,且维持时间较长,接种后60天仍维持在1:26以上,能很好地抵抗NDV强毒的攻击。NDV还可以和其他病原混合制备成灭活联苗,彼此间不会出现相互干扰作用。
④免疫抑制病 一些能够产生免疫抑制的病原,可不同程度地降低ND疫苗的免疫效果。Giambrone等(1986,1990)的研究表明,蛋鸡和肉鸡出壳后饲养在有传染性法氏囊病毒(IBDV)污染的环境里,会出现对ND疫苗接种的免疫抑制,表现为ND HI抗体效价降低。28日龄时免疫ND,35日龄时试验组鸡(有IBDV污染)HI抗体滴度(GMT)比对照组鸡(无IBDV污染)显著降低。鸡传染性贫血因子引起的免疫抑制,可导致灭活的新城疫疫苗二次免疫失败。具有免疫抑制作用的病原还有:传染性法氏囊病毒、马立克氏病毒、淋巴白血病病毒、网状内皮增生症病毒、呼肠孤病毒、传染性贫血因子等。
⑤霉菌毒素的影响 发霉的饲料中含有黄曲霉或棕曲霉毒素时,只需要很小的量就能对鸡的免疫系统产生抑制作用,从而使ND疫苗免疫鸡的HI效价达不到所要求的水平。
⑥日龄的影响 雏鸡的免疫器官发育不健全,40日龄前主要靠法氏囊的B细胞产生的体液免疫,40日龄后T细胞才参与免疫应答,70日龄后鸡的免疫器官才发育成熟。因此,雏鸡所表现的免疫应答不如成鸡。
3. 减少应激,提高动物机体健康水平,提高免疫效果
动物的机体状况和健康水平直接影响动物免疫的质量。动物机体的免疫功能在一定程度上受到神经、体液和内分泌的调节,在环境过冷过热、湿度过大、通风不良、拥挤、饲料突然改变、运输、转群等应激因素的影响下,机体肾上腺皮质激素分泌增加。肾上腺皮质激素能显著损伤T淋巴细胞,对巨噬细胞也有抑制作用,增加IgG的分解代谢。所以,当动物处于应激反应敏感期时接种疫苗,就会减弱动物的免疫能力。此外,维生素及许多其它养分都对动物免疫力有显著影响。养分缺乏,特别是缺乏维生素A、D、B、E和多种微量元素及全价蛋白时能影响机体对抗原的免疫应答,免疫反应明显受到抑制。以鸡为例,试验表明,雏鸡断水、断食48小时,法氏囊、胸腺和脾脏重量明显下降,脾脏内淋巴细胞数减少,网状内皮系统细菌清除率降低,即机体免疫能力下降。
4. 建立免疫监测制度 主要是通过定期抽样检查鸡群的抗体水平(HI抗体),不仅可帮助建立科学的免疫程序,而且可以判定和预见疾病的发生、发展。其中,HI抗体水平的高低,可反应鸡群是否能够抵抗野毒感染,是否需要加强免疫等。有条件的鸡场,应建立免疫监测手段,定期对鸡群抽样采取血清作血凝抑制试验。实验证明,母鸡血清中的HI价与当时所产种蛋的卵黄HI价以及用此种鸡蛋孵出的雏鸡的血清HI价,二者大致相近,故可通过检测卵黄HI价来了解母鸡和雏鸡血清中的抗体水平,从而确定接种疫苗的时机。监测的鸡数应根据鸡群的大小而定:1000只以下时,不少于10~20只;5000只以下时,不少于30~50只;10000只以下时,不少于50~100只。商品肉鸡应该在1:27左右、蛋鸡和种鸡应在1:29以上时,才能够很好的保护;若抗体突然上升很高,达到1:213以上,表明很有可能发生了野毒感染。
5. 注重生物安全 疫苗免疫是防制ND的一种重要方法,但仅以疫苗免疫为控制策略而忽视整个生物安全体系的建立是极其错误的。疫苗免疫只可能减少NDV可能侵入鸡群而带来的经济损失;尽可能降低发病几率,而决不能阻止NDV进入鸡群,更不能消灭鸡群内已经存在的病毒。鸡群如果暴露于强病毒包围的环境中,感染率是极高的,即使是免疫鸡群,感染和发病也是完全可能的。作为群体免疫应答这一生物学系统,即使是频繁接种各种ND疫苗。也不可能产生100%的保护。因为群体中每一个体的免疫应答水平是不相同的,病毒与鸡群之间的相互作用复杂而微妙,出现的结果不一。强毒株感染鸡群后,产生良好免疫的鸡不发病,这应该占大多数,免疫不良者或说免疫不确实的鸡势必出现非典型性ND症状。不可忽视的一个问题是这两种情况下,鸡本身是带毒的,后者还是排毒者。又由于个体免疫的差异,现场强毒的毒力和数量与鸡的免疫力之间不断的相互作用,逐渐发生复杂的生物学量和质的变化,在一定条件下,强毒就有可能突破某些免疫力不足的鸡的免疫保护,在体内复制增殖,使鸡出现ND典型症状和病变。
正因为如此,建立健全的生物安全体系至关重要。首先,要建立科学的严格的卫生防疫制度和措施,诸如入场人员的淋浴更衣消毒,车辆用具物品带入生产区时先消毒处理,鸡场生活和生产区分开,生产区内净、污道分开,粪便垫料及污水的处理,场区和鸡舍的消毒,鼠兽的预防,生产区工具专用,谢绝无关人员参观等各项措施真正落实到位,以切断传播途径。其次,要实行场区或栋舍化的全进全出制度,以彻底消除NDV在鸡场内传播。第三鸡舍在进鸡前必须彻底清扫、冲刷、消毒,并有适当的空闲期,使鸡舍得以净化,进鸡前至少消毒2次。第四,要改变生产模式,关注鸡的福利。
6.关注新城疫病毒的变异
近几年来,在许多免疫过的鸡场甚至搞抗体水平的鸡场,常有免疫失败的发生,而且均有较为明显的新城疫症状和病变,和以往的非典型新城疫有较大的不同,然而通过基因分型发现,造成这一现象的大部分病毒属于基因Ⅶ,许多人担心原先的疫苗(属于基因Ⅰ、Ⅱ、Ⅲ、Ⅳ型)是否能保护鸡抵抗基因Ⅶ的强毒。通过大量实验表明,虽然不同基因型之间有一定抗原差异,但依然能够保护或者部分保护。
扬州大学应用基因VII型Y98毒株和T99毒株、NDV F48E8毒株、LaSota毒株和NDV分离株为基因为III和II型,分别制备油乳剂灭活苗,各免疫4日龄肉鸡50只,免疫后3~5周分别用Y98毒株和T99毒株攻击。结果:Y98毒株免疫组为98%(49/50),而LaSota毒株和NDV分离株组均为0,NDV F48E8为10%(5/50);T99攻击组:T99毒株免疫组为98%(49/50),Y98毒株免疫组为96%(48/50),而LaSota毒株组为0,NDV F48E8为10%(5/50)。Yu L等(2001)进行疫苗株LaSota与田间分离株CH-A7/96之间的交叉保护试验。CH-A7/96油乳剂疫苗免疫鸡能完全抗住CH-A7/96及CH-F48E9/44的攻击,不发病,无死亡;LaSota疫苗株免疫鸡,虽以CH-F48E9/44攻毒时有90%的保护率,但用CH-A7/96攻毒时的保护率仅只有40%。田夫林等(2003)对11株NDV分离株作了致病性和抗原性变异的研究,它们中HI值存在明显差异,Liu株血清抗体对C30、Lan、F48株抑制价低3~4个滴度,H株血清抗体与C30、Lan、F48株的抑制价也低了3~4个滴度,C30对F48、Liu的抑制价低2个滴度,说明目前分离株与F48、早期分离株和疫苗株在凝集原上有明显差异。刘文斌等(2005)对经属基因Ⅱ型LaSota、Clone30免疫具高抗体的42日龄鸡用基因Ⅶ型JL-Ⅱ分离株、基因Ⅸ型的F48E9攻击,JL-Ⅱ致死率达28.5%,F48E9的致死率7.14%,表明生产中常用的疫苗并不能完全保护目前流行株的攻击。秦卓明等人(2007)通过17株新城疫病毒的HI交叉抑制试验、鸡胚中和试验和细胞中和试验证实:尽管三者使用的毒株不尽相同,但结论基本一致:
针对疫区生产中存在的问题,建议如下:首先,坚持灭活苗和弱毒苗的联合免疫,提高鸡群体液抗体水平,种鸡抗体保持在1:210以上,特别是开产前的种鸡和蛋鸡,最好利用ND灭活苗单独免疫,以后每月,弱毒苗免疫1次,每3个月免疫1次灭活苗;其次,有条件的鸡场可考虑利用流行株进行自家苗免疫。第三,加强管理,提高机体健康水平。
(三)发生新城疫的措施
鸡群一旦发生可疑新城疫,首先应采取隔离措施,防止疫情扩大。同时报告兽医检查,经确诊后,及时报告当地政府,划定疫区进行封锁;其次应根据鸡群不同的日龄采取相应的措施。雏鸡可采用2~3倍量的Ⅳ系或Clone30紧急接种,2月龄以上的鸡可使用2~3倍量的Ⅰ系或克隆Ⅰ系对鸡群进行紧急接种,开产的种鸡可接种流行株灭活苗或使用2~3倍量的克隆Ⅰ系紧急接种,接种顺序为假定健康群,然后为可疑群,最后为病鸡群;实践证明:紧急接种,能够减少一部分死亡和缩短病程,减少生产损失。高免血清和卵黄抗体有一定作用,但成本太高,且存在很多隐患,一般不建议。第三,采取紧急消毒措施,对鸡舍、用具等使用高浓度的“驰骋一片净”、2%烧碱溶液进行消毒;对病死鸡尸体、内脏及排泄物等应进行焚烧或深埋,或是严格消毒处理。
参考文献
[1]甘孟候 主编.中国禽病学.第一版. 北京:中国农业出版社,1997.
[2] 翟文栋, 鸭新城疫强毒的分离鉴定. 中国家禽,2007,28(6):4-6.
[3]秦卓明,何叶峰,徐怀英,等.种鸡感染新城疫强毒的诊断及其防制. 山东家禽,2002,5:35-37.
[4]Saif YM 主编. 禽病学.苏敬良,高福,索勋主译.第十一版.北京:中国农业出版社,2005.
[5]谭雷涛,秦卓明,马保臣. 一株新城疫重组强毒的分离鉴定与分子特性分析.畜牧兽医学报,2007,11:1217-1223
[6]秦卓明,马保臣,袁小远.新城疫分离毒HN基因的分子特性和片段同源相关性病毒学报.2007,1:39-45
[7]马保臣;何叶峰;徐怀英;秦卓明.高抗体水平种鸡NDV的分离鉴定和分子特性.微生物学通报.2007,1:95-100
[8]世界动物卫生组织(OIE)著.哺乳动物、禽、蜜蜂A和B类疫病诊断试验和疫苗标准手册.农业部畜牧兽医局译.第一版.北京:中国农业科学技术出版社,2002.
[9]Aldous EW, Alexander DJ. Detection and differentiation of
[10]Runa Panda, Zhuhui Huang, Subbiah Elankumaran, et al. Role of fusion protein cleavage site in the virulence of
[11]Nagai Y, Klenk HD, Rott R. Proteolytic cleavage of the viral glycoproteins and its significance for the virulence of
[12]Peeters BP, De Leeuw OS, Koch G, et al. Rescue of Newcastle disease virus from cloned cDNA: evidence that cleavability of the fusion protein is a major determinant for virulence. J Vir,1999, 73(6): 5001-5009.
[13]Ballagi-Pordany A, Wehman E, Herczeg J, et al. Identification and grouping of
[14] Toyoda T, Sakaguchi T, Hirota H, et al.
[15] Liu XF, Wan.HQ, Ni XX, et al. Pathotypical and genotypical characterization of strains of Newcastle Disease virus isolated from outbreaks in chicken and goose in some regions of China during 1985~2001. Arch Virol,2003,148(7):1387-1403.
[16]Takehara K, Shinomiya T, Kobayashi H. et al. Characterization of
[17]Shengqing Y, Kishida N, Ito H, et al. Generation of velogenic
[18] Liu X F, Wan H Q, Ni X X, Wu Y T and Liu W B.Pathotypical and genotypical characterization of strains of Newcastle Disease virus isolated from outbreaks in chicken and goose in some regions of China during 1985~2001[J].Archives of Virology,2003,148(7):1387~1403.
[19]Qin ZM, Tan LT, Xu HY, Ma BC, Wang YL, Yuan XY, Liu WJ.Pathotypical characterization and molecular epidemiology of Newcastle disease virus isolates from different hosts in China from 1996 to 2005. J Clin Microbiol. 2008 Feb;46(2):601-11
[20]Qin Z, Sun L, Ma B, Cui Z, Zhu Y, Kitamura Y, Liu W. F gene recombination between genotype II and VII Newcastle disease virus. Virus Res. 2008 Feb;131(2):299-303.
鸡鸭鹅病防治网 为规模化养殖企业(场)提供疫病防控最佳解决方案!!!http://gallops99.blog.163.com/
禽病实战专家咨询:13939028068 (技术总监-魏老师 个人 微信:CC-zbs 欢迎交流)博客:http://blog.sina.com.cn/gallopsbj
养殖安880、驰骋钙、蚊蝇一喷净等产品销售顾问:成龙在路上
13838248089 博客: http://lyrswcl.blog.163.com



发表评论共有0访客发表了评论
暂无评论,快来抢沙发吧!